Principles
of Classification
Think about an elephant. Develop
a mental image of it. How would you describe it to someone who has never
seen one? Take a moment to consider carefully . . .
Click
the button to see if
your mental image was accurate.

Very likely your mental image was a visual
one like the picture. Humans primarily emphasize traits that can be seen
with their eyes since they mostly rely on their sense of vision.
However, there is no reason that an elephant or any other organism could not
be described in terms of touch, smell, and/or sound as well. Think about
an elephant again but this time in terms of non-visual traits . . .
Not surprisingly, biologists also classify
organisms into different categories mostly by judging degrees of apparent
similarity and difference that they can see. The assumption is that the
greater the degree of physical similarity, the closer the biological
relationship.
On discovering an unknown organism,
researchers begin their classification by looking for anatomical features that
appear to have the same function as those found on other species. The
next step is determining whether or not the similarities are due to an
independent evolutionary development or to descent from a common ancestor.
If the latter is the case, then the two species are probably closely related
and should be classified into the same or near biological categories.
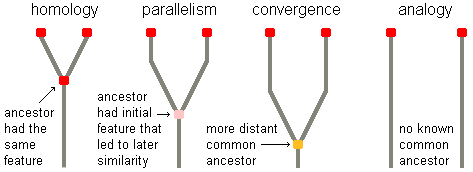
Homologies
are anatomical features, of different organisms, that have a similar
appearance or function because they were inherited from a common ancestor that
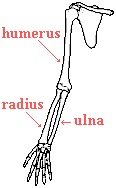
also had them. For instance, the forelimb of a bear, the wing of a bird,
and your arm have the same functional types of bones as did our shared
reptilian ancestor. Therefore, these bones are
homologous structures. The more homologies two organisms possess, the
more likely it is that they have a close genetic relationship.
There can also be non-homologous structural
similarities between species. In these cases, the common ancestor did
not have the same anatomical structures as its descendants. Instead, the
similarities are due to independent development in the now separate
evolutionary lines. Such misleading similarities are called homoplasies.
Homoplastic structures can be the result of parallelism, convergence,
analogies, or mere chance.
Parallelism,
or parallel evolution, is a similar evolutionary development in different
species lines after divergence from a common ancestor that
did not have the characteristic but did have an initial anatomical
feature that led to it. For instance, some South American and African
monkeys evolved relatively large body sizes independently of each other.
Their common ancestor was a much smaller monkey but was otherwise reminiscent
of the later descendant species. Apparently, nature selected for larger
monkey bodies on both continents during the last 30 million years.
Convergence,
or convergent evolution, is the development of a similar anatomical feature in
distinct species lines after divergence from a common ancestor that did not
have the initial trait that led to it. The common ancestor is
usually more distant in time than is the case with parallelism.
The similar appearance and predatory behavior of North American wolves and
Tasmanian wolves is an example. The former is a placental
mammal and the latter is an Australian marsupial.
Their common ancestor lived during the age of the dinosaurs more than
100 million years ago and was very different from these descendants today.
There are, in fact, a number of other Australian marsupials that are striking
examples of convergent evolution with placental mammals elsewhere.
 |
|
 |
|
Tasmanian
wolf or tiger
(now extinct)
|
|
Mexican
wolf
|
 Examples
of Convergent Evolution--ant eating
mammals from four continents
Examples
of Convergent Evolution--ant eating
mammals from four continents
This link takes
you to an external website. To return here, you must click
the "back"
button on your browser
program. |
Both parallelism and convergence are
thought to be due primarily to separate species lines experiencing the same
kinds of natural selection pressures over long periods of time.
Analogies
are anatomical features that have the same form or function in different
species that have no known common ancestor. For instance, the wings of a
bird and a butterfly are analogous structures because they are superficially
similar in shape and function. Both of these very distinct species lines
solved the problem of getting off of the ground in essentially the same way.
However, their wings are quite different on the inside. Bird wings have
an internal framework consisting of bones, while butterfly wings do not have
any bones at all and are kept rigid mostly through fluid pressure.
Analogies may be due to homologies or homoplasies, but the common ancestor, if
any, is unknown.
Problems in Classifying
Organisms
Listing characteristics that distinguish
one species from another has the effect of making it appear that the species
and their distinctive attributes are fixed and eternal. We must always
keep in mind that they were brought about by evolutionary processes that
operated not merely at some time in the distant past, but which continue to
operate in the present and can be expected to give rise to new forms in the
future. Species are always changing. As a
consequence, they are essentially only a somewhat arbitrarily defined point
along an evolutionary line.
It is also important to realize that most
species are physically and genetically diverse.
Many are far more varied than humans. When you think of an
animal, such as the jaguar shown here, and describe it in terms of its
specific traits (fur color patterns, body shape, etc.), it is natural to
generalize and to think of all jaguars that way. To do so, however, is
to ignore the reality of diversity in nature.
Another problem in classifying a newly
discovered organism is in determining the specific characteristics that
actually distinguish it from all other types of organisms. There is
always a lively debate among researchers over defining new species because it
is not obvious what are the most important traits. There
are two schools of thought in resolving this dilemma. The first defines
new species based on minor differences between organisms. This is the splitter
approach. The second tends to ignore minor differences and to
emphasize major similarities. This lumper
approach results in fewer species being defined.
Ideally, this dispute could be settled by
breeding experiments--if two organisms can mate and produce fertile offspring,
they are probably members of the same species. However, we must be
careful because members of very closely related species can sometimes produce
offspring together, and a small fraction of those may be fertile. This
is the case with mules, which are the product of mating between female
horses and male donkeys. About one out
of 10,000 mules is fertile. Does this mean that
horses and donkeys are in the same species? Whatever the answer may be,
it is clear that species are not absolutely distinct entities, though by
naming them, we implicitly convey the idea that they are.
Tigons
and Ligers--what happens when tigers and
lions mate
This link takes you
to an external website. To return here, you must
click
the "back" button on your
browser program. |
Breeding
experiments are rarely undertaken to determine species boundaries because of
the practical difficulties. It is time consuming and wild animals do not
always cooperate. Using this kind of reproductive data for defining
species from the fossil record is impossible since we cannot go back in time
to observe breeding patterns and results. Comparisons of DNA
sequences are now becoming more commonly used as an aid in distinguishing
species. If two animals share a great many DNA sequences, it is likely
that they are at least closely related. Unfortunately, this usually does
not conclusively tell us that they are members of the same species.
Therefore, we are still left with morphological
characteristics as the most commonly used criteria for identifying species
differences.
The Linnaean scheme for classification of
living things lumps organisms together based on presumed homologies. The
assumption is that the more homologies two organisms share, the closer they
must be in terms of evolutionary distance. The higher,
more inclusive divisions of the Linnaean system are created by including
together closely related clusters of the immediately lower divisions.
The result is a hierarchical system
of classification with the highest category consisting of all living things.
The lowest category consists of a single species. Each of the categories
above species can have numerous subcategories. In
the example below, only two genera (plural of genus) are
listed per family but there could be many more or only one.
| order |
| family |
family |
| genus |
genus |
genus |
genus |
| species |
species |
species |
species |
species |
species |
species |
species |
Most researchers today
take a cladistics approach to
classification. This involves making a distinction between derived
and primitive traits when evaluating the
importance of homologies in determining placement of organisms within the
Linnaean classification system. Derived traits are those that have
changed from
the ancestral form and/or function. An example is the foot of a modern
horse. Its distant early mammal ancestor had five digits. The
bones of these digits have been largely fused together in horses giving them
essentially only one toe with a hoof. In contrast, primates have
retained the primitive characteristic of having five digits on the ends of
their hands and feet. Animals sharing a great many homologies that
were recently derived, rather than only ancestral, are more likely to have a
recent common ancestor. This assumption is the basis of cladistics.
Next
Topic
Previous
Topic
Main Menu