Animal Kingdom Unit
Squid Dissection Lab
As part of our continued exploration of the animal kingdom today we will perform a dissection of a common European squid, which is a member of the Class Cephalopoda within the Phylum Mollusk. This lab should introduce you to several of the major tissue types and organ systems found within these animals.
If you were absent and missed the lab click HERE for a video of the squid dissection.
Squid Phylogeny
| Kingdom | Animalia |
| Phylum | Mollusca |
| Class | Cephalopoda |
| Order | Teuthida |
| Family | Loliginidae |
| Genus | Loligo |
| Species | vulgaris |
Therefore the scientific name of the squid you will dissect is: Loligo vulgaris
Evolution
Crown coleoids (the ancestors of octopuses and squid) diverged at the end of the Paleozoic, in the Permian. Squid diverged during the Jurassic, but many squid families appeared in or after the Cretaceous. Both the coleoids and the teleost fish were involved in much adaptive radiation at this time, and the two modern groups resemble each other in size, ecology, habitat, morphology and behaviour, however some fish moved into fresh water while the coleoids remained in marine environments.
The ancestral coleoid was probably nautiloid-like with a strait septate shell that became immersed in the mantle and was used for buoyancy control. Four lines diverged from this, Spirulida (with one living member), the cuttlefishes, the squids and the octopuses. Squid have differentiated from the ancestral mollusc such that the body plan has been condensed antero-posteriorly and extended dorso-ventrally. What may have been the foot of the ancestor is modified into a complex set of appendages around the mouth. The sense organs are highly developed and include advanced eyes similar to those of vertebrates.
The ancestral shell has been lost, with only an internal gladius, or pen, remaining. The pen, made of a chitin-like material, is a feather-shaped internal structure that supports the squid's mantle and serves as a site for muscle attachment. The cuttlebone or sepion of the Sepiidae is calcareous and appears to have evolved afresh in the Tertiary.
Description
Squid are soft-bodied molluscs whose forms evolved to adopt an active predatory lifestyle. The head and foot of the squid are at one end of a long body, and this end is functionally anterior, leading the animal as it moves through the water. A set of eight arms and two distinctive tentacles surround the mouth; each appendage takes the form of a muscular hydrostat and is flexible and prehensile, usually bearing disc-like suckers.
The suckers may lie directly on the arm or be stalked. Their rims are stiffened with chitin and may contain minute toothlike denticles. These features, as well as strong musculature, and a small ganglion beneath each sucker to allow individual control, provide a very powerful adhesion to grip prey. Hooks are present on the arms and tentacles in some species, but their function is unclear. The two tentacles are much longer than the arms and are retractile. Suckers are limited to the spatulate tip of the tentacle, known as the manus.
In the mature male, the outer half of one of the left arms is hectocotylised – and ends in a copulatory pad rather than suckers. This is used for depositing a spermatophore inside the mantle cavity of a female. A ventral part of the foot has been converted into a funnel through which water exits the mantle cavity.
The main body mass is enclosed in the mantle, which has a swimming fin along each side. These fins are not the main source of locomotion in most species. The mantle wall is heavily muscled and internal. The visceral mass, which is covered by a thin, membranous epidermis, forms a cone-shaped posterior region known as the "visceral hump". The mollusc shell is reduced to an internal, longitudinal chitinous "pen" in the functionally dorsal part of the animal; the pen acts to stiffen the squid and provides attachments for muscles.
On the functionally ventral part of the body is an opening to the mantle cavity, which contains the gills (ctenidia) and openings from the excretory, digestive and reproductive systems. An inhalant siphon behind the funnel draws water into the mantel cavity via a valve. The squid uses the funnel for locomotion via precise jet propulsion. In this form of locomotion, water is sucked into the mantle cavity and expelled out of the funnel in a fast, strong jet. The direction of travel is varied by the orientation of the funnel. Squid are strong swimmers and certain species can "fly" for short distances out of the water.
Camouflage
Squid make use of different kinds of camouflage, namely active camouflage for background matching (in shallow water) and counter-illumination. This helps to protect them from their predators and allows them to approach their prey.
The skin is covered in controllable chromatophores of different colours, enabling the squid to match its coloration to its surroundings. The play of colours may in addition distract prey from the squid's approaching tentacles. The skin also contains light reflectors called iridophores and leucophores that, when activated, in milliseconds create changeable skin patterns of polarized light. Such skin camouflage may serve various functions, such as communication with nearby squid, prey detection, navigation, and orientation during hunting or seeking shelter. Neural control of the iridophores enabling rapid changes in skin iridescence appears to be regulated by a cholinergic process affecting reflectin proteins.
Some mesopelagic squid such as the firefly squid (Watasenia scintillans) and the midwater squid (Abralia veranyi) use counter-illumination camouflage, generating light to match the downwelling light from the ocean surface. This creates the effect of countershading, making the underside lighter than the upperside.
Counter-illumination is also used by the Hawaiian bobtail squid (Euprymna scolopes), which has symbiotic bacteria (Aliivibrio fischeri) that produce light to help the squid avoid nocturnal predators. This light shines through the squid's skin on its underside and is generated by a large and complex two-lobed light organ inside the squid's mantle cavity. From there, it escapes downwards, some of it travelling directly, some coming off a reflector at the top of the organ (dorsal side). Below there is a kind of iris, which has branches (diverticula) of its ink sac, with a lens below that; both the reflector and lens are derived from mesoderm. The squid controls light production by changing the shape of its iris or adjusting the strength of yellow filters on its underside, which presumably change the balance of wavelengths emitted. Light production shows a correlation with intensity of down-welling light, but it is about one third as bright; the squid can track repeated changes in brightness. Because the Hawaiian bobtail squid hides in sand during the day to avoid predators, it does not use counter-illumination during daylight hours.
Controllable chromatophores of different colours in the skin
of a squid allow it to change its coloration and patterns rapidly, whether for
camouflage or signalling.
Predator distraction with ink
Fossil Loligosepia aalensis from the lower Jurassic; the ink
sac is still full of black eumelanin pigment
Squid distract attacking predators by ejecting a cloud of ink, giving themselves an opportunity to escape. The ink gland and its associated ink sac empties into the rectum close to the anus, allowing the squid to rapidly discharge black ink into the mantle cavity and surrounding water.The ink is a suspension of melanin particles and quickly disperses to form a dark cloud that obscures the escape manoeuvres of the squid. Predatory fish may also be deterred by the alkaloid nature of the discharge which may interfere with their chemoreceptors.
Behavior
Locomotion
Squid can move about in several different ways. Slow movement is achieved by a gentle undulation of the muscular lateral fins on either side of the trunk which drives the animal forward. A more common means of locomotion providing sustained movement is achieved using jetting, during which contraction of the muscular wall of the mantle cavity provides jet propulsion.
Slow jetting is used for ordinary locomotion, and ventilation of the gills is achieved at the same time. The circular muscles in the mantle wall contract; this causes the inhalant valve to close, the exhalant valve to open and the mantle edge to lock tightly around the head. Water is forced out through the funnel which is pointed in the opposite direction to the required direction of travel. The inhalant phase is initiated by the relaxation of the circular muscles causes them to stretch, the connective tissue in the mantle wall recoils elastically, the mantle cavity expands causing the inhalant valve to open, the exhalant valve to close and water to flow into the cavity. This cycle of exhalation and inhalation is repeated to provide continuous locomotion.
Fast jetting is an escape response. In this form of locomotion, radial muscles in the mantle wall are involved as well as circular ones, making it possible to hyper-inflate the mantle cavity with a larger volume of water than during slow jetting. On contraction, water flows out with great force, the funnel always being pointed anteriorly, and travel is backwards. During this means of locomotion, some squid exit the water in a similar way to flying fish, gliding through the air for up to 50 m (160 ft), and occasionally ending up on the decks of ships.
Feeding
Squid are carnivores, and, with their strong arms and suckers, can overwhelm relatively large animals efficiently. Prey is identified by sight or by touch, grabbed by the tentacles which can be shot out with great rapidity, brought back to within reach of the arms, and held by the hooks and suckers on their surface. In some species, the squid's saliva contains toxins which act to subdue the prey. These are injected into its bloodstream when the prey is bitten, along with vasodilators and chemicals to stimulate the heart, and quickly circulate to all parts of its body. The deep sea squid Taningia danae has been filmed releasing blinding flashes of light from large photophores on its arms to illuminate and disorientate potential prey.
Although squid can catch large prey, the mouth is relatively small, and the food must be cut into pieces by the chitinous beak with its powerful muscles before being swallowed. The radula is located in the buccal cavity and has multiple rows of tiny teeth that draw the food backwards and grind it in pieces. The deep sea squid Mastigoteuthis has the whole length of its whip-like tentacles covered with tiny suckers; it probably catches small organisms in the same way that flypaper traps flies. The tentacles of some bathypelagic squids bear photophores which may bring food within its reach by attracting prey.
Squid are among the most intelligent invertebrates. For example, groups of Humboldt squid hunt cooperatively, spiralling up through the water at night and coordinating their vertical and horizontal movements while foraging.
Reproduction
Courtship in squid takes place in the open water and involves the male selecting a female, the female responding, and the transfer by the male of spermatophores to the female. In many instances, the male may display to identify himself to the female and drive off any potential competitors. Elaborate changes in body patterning take place in some species in both agonistic and courtship behaviour. The Caribbean reef squid (Sepioteuthis sepioidea), for example, employs a complex array of colour changes during courtship and social interactions and has a range of about 16 body patterns in its repertoire.
The pair adopt a head-to-head position, and "jaw locking" may take place, in a similar manner to that adopted by some cichlid fish. The heterodactylus of the male is used to transfer the spermatophore and deposit it in the female's mantle cavity in the position appropriate for the species; this may be adjacent to the gonopore or in a seminal receptacle.
Squid eggs
The sperm may be used immediately or may be stored. As the eggs pass down the oviduct, they are wrapped in a gelatinous coating, before continuing to the mantle cavity, where they are fertilised. In Loligo, further coatings are added by the nidimental glands in the walls of the cavity and the eggs leave through a funnel formed by the arms. The female attaches them to the substrate in strings or groups, the coating layers swelling and hardening after contact with sea water. Loligo sometimes forms breeding aggregations which may create a "community pile" of egg strings. Some pelagic and deep sea squid do not attach their egg masses, which float freely.
Ecology
Squid mostly have an annual life cycle, growing fast and dying soon after spawning. The diet changes as they grow but mostly consists of large zooplankton and small nekton. In Antarctica for example, krill is the main constituent of the diet, with other food items being amphipods, other small crustaceans, and large arrow worms. Fish are also eaten, and some squid are cannibalistic.
As well as occupying a key role in the food chain, squid are an important prey for predators including sharks, sea birds, seals and whales. Juvenile squid provide part of the diet for worms and small fish. When researchers studied the contents of the stomachs of elephant seals in South Georgia, they found 96% squid by weight. In a single day, a sperm whale can eat 700 to 800 squid, and a Risso's dolphin entangled in a net in the Mediterranean was found to have eaten angel clubhook squid, umbrella squid, reverse jewel squid and European flying squid, all identifiable from their indigestible beaks. Ornithoteuthis volatilis, a common squid from the tropical Indo-Pacific, is predated by yellowfin tuna, longnose lancetfish, common dolphinfish and swordfish, the tiger shark, the scalloped hammerhead shark and the smooth hammerhead shark. Sperm whales also hunt this species extensively as does the brown fur seal. In the Southern Ocean, penguins and wandering albatrosses are major predators of Gonatus antarcticus.
Introduction to the Mollusk Body Organization
Mollusks are soft-bodied animals, more familiarly known as clams, octopods, and snails. They are bilaterally symmetrical, with well-developed digestive, circulatory, excretory, and respiratory systems. A calcareous shell may (e.g., snails, clams) or may not (e.g., slugs, octopods) be present.
Mollusks are closely related to annelids (segmented worms) in their mode of development and type of larva, but they are distinguished from the latter by a lack of segmentation. In addition, the extensive coelom, so important in annelid locomotion, is reduced in size in most mollusks; it is usually restricted to the area surrounding the heart and to spaces within the gonads and kidneys. Most mollusks are slow-moving, creeping animals, but in some instances the organization of the body regions has been modified so that swift movements are possible.
In an evolutionary sense, mollusks are extremely plastic, and they demonstrate highly successful adaptations to a variety of habitats. The considerable adaptive radiation exhibited by members of this phylum is reflected in the varied functional morphology of the group. In numbers of species, they are among the most abundant of all organisms. Although primarily marine, representatives are found in fresh water and on land.
All mollusks can theoretically be derived from a generalized body plan consisting of three main body regions: head-foot, visceral mass, and mantle. The head-foot region is the locomotory and sensory portion of the body, upon which rides the visceral mass containing the excretory, digestive, and circulatory organs. The third component of the body, the mantle, forms a fleshy cover over the visceral mass and a skirt around the foot. It secretes the shell ( in those mollusks that have one), which lies on its outer surface, contains the gills (ctenidia), and itself may function in respiration. The mantle alone might be considered the hallmark of the mollusks, for the flexibility of the mantle in both form and function has contributed greatly to the success of the group.
Dissection of the Squid
Click HERE for a Virtual Squid Dissection Lab
Use this Virtual Squid Dissection Lab as a guide to help you dissect your specimen.
External Anatomy
As the name indicates (cephalo = head; pod = foot), the head-foot is dominant in cephalopods. These are swift-moving carnivores, and their bodies are quite streamlined. The shell is either not well developed or entirely absent. When present, the shell is internal, except in the most ancient lineages (e.g., Nautilus).
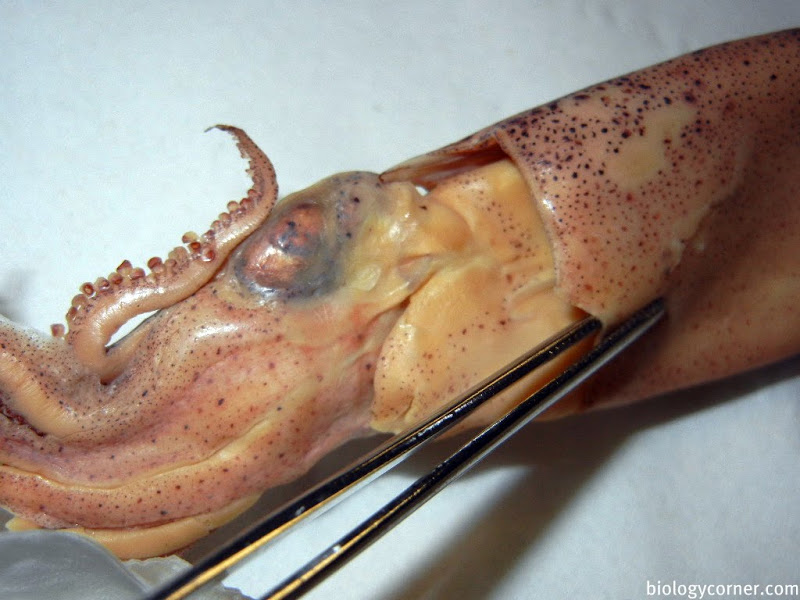
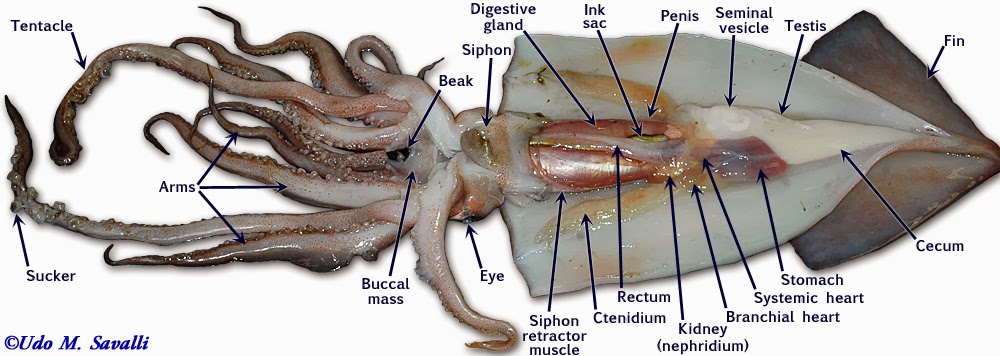
Examine a preserved specimen of the squid. Note that there is no external shell and that the major part of the body is enclosed in a soft, fleshy mantle and is sharply demarcated from the rest of the squid body by the collar. Opposite the pointed end of the animal (the apex), you will find the head-foot region. The eight arms and two longer tentacles are derivatives of the foot and surround the mouth. Turn back the arms and tentacles to reveal a muscular membrane running from their bases to the mouth. This membrane is composed of an outer lobed buccal membrane with suckers on its inner surface and an inner peristomial membrane. Notice the horny beak protruding from the mouth. Locate the large, well-developed eyes and the fold of tissue behind the eye, called the olfactory crest.
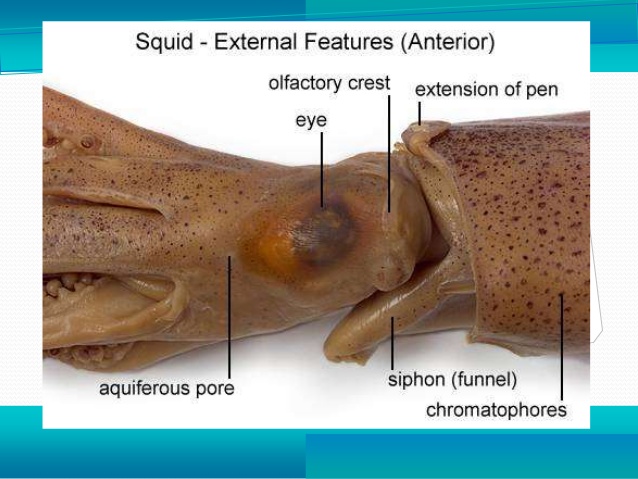
Orientation at this point may cause problems, so align your specimen in such a way that the following descriptions will not be confusing. Place the animal so the apex is furthest from you and the arms are closest to you. Turn the animal so that the siphon (the tubular projections between the collar and the head-foot region) is facing you. It too is a derivative of the foot. The apex, which bears the visceral mass (digestive organs), is dorsal and the arms are ventral. The surface facing you is posterior. The eyes are on the left and right sides of the body, and the head structures have moved to a position that is dorsal to the foot. Contrary to popular belief, the squid moves in the direction of its tentacles and arms. The fin is the trailing end of the squid when it is motion and acts like a rudder to help them steer to change right and left directional movements.

Internal Anatomy
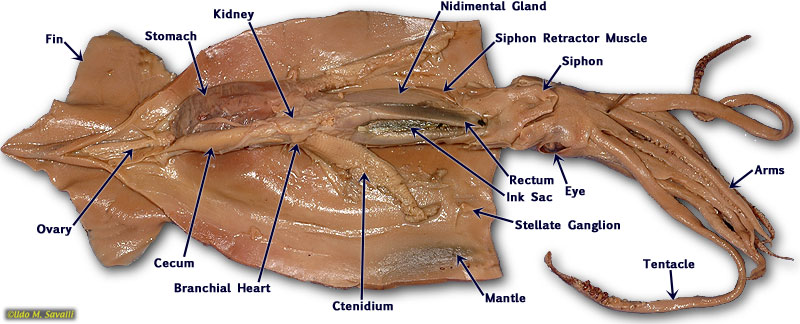
The mantle is a fleshy cover that incompletely surrounds most of the organs of the body. The space between the fleshy mantle lobes is called the mantle cavity. The openings of the digestive, excretory, and reproductive systems are all found within the mantle cavity. The mantle also functions in secretion of the shell. However, the most distinctive function of the mantle cavity is to provide a space for housing the delicate ctenidia (gills), thus protecting them from the hazards of the environment and permitting an oriented flow of water across them.
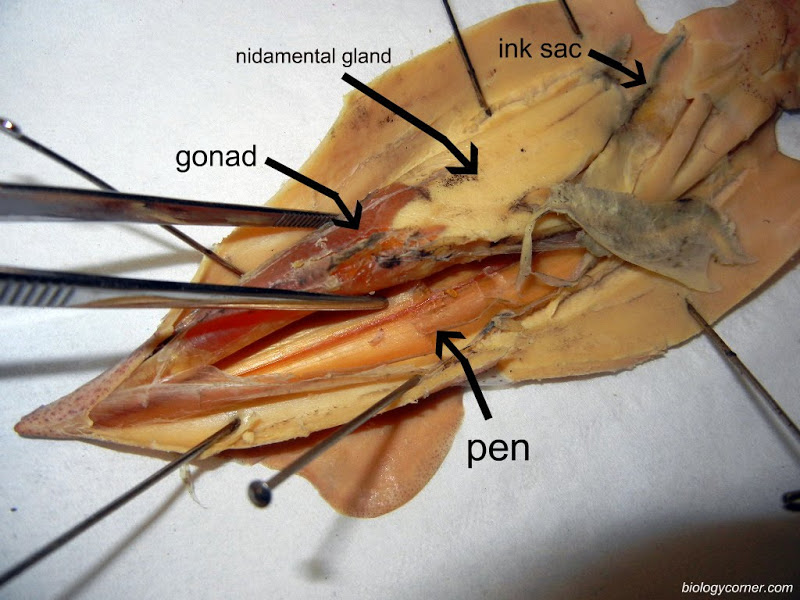
Primitive cephalopods show a chambered shell (e.g., Nautilus), but in others the shell is internal or absent. In Loligo, the chitinous endoskeletal element is known as the pen. You will be able to feel the stiff pen running the full length of your animal under the dorsal anterior surface. Of what advantage might a reduced or internal shell be to cephalopods?
The most obvious organs of the mantle are the paired ciliated gills or ctenidia. Strictly speaking, a ctenidium is a respiratory structure that includes ciliated filaments. The ciliary tracts on the filaments draw respiratory water currents into the mantle cavity on either side. The two currents filter through the ctenidial filaments, meet dorsally, and pass backward as an exhalent stream out of the mantle cavity. Gland cells on the filaments secrete mucus, and particles brought into the mantle cavity with the respiratory current are filtered out. Tracts of cilia on the ctenidia direct rejected material toward the midline.
Cephalopods are active predators, streamlined for quick motion; they therefore require a streamlining of their respiratory current. The walls of the mantle cavity are highly muscularized, and movement of water within the cavity is no longer dependent on ciliary action. With the ventral side of the squid facing toward you, observe the construction of the mantle collar and its relationship to the siphon. The siphon is derived from the foot. In life, the mantle cavity expands by muscular action, water enters, and then the collar locks tightly against the head, leaving the siphon as the only exit pathway from the mantle cavity. The siphon is well equipped with muscles and can be pointed for making directed jet-propulsive movements.
Open the mantle cavity by making an incision that runs the entire length of the posterior surface from siphon to apex. Dissect with care so as not to disturb the internal organs. Notice that the mantle consists of a thick layer of circular muscles surrounded by integument, with dorsolateral muscularized extensions, the fins. Turn the cut mantle edges laterally and pin them out to expose the internal organs clearly. Note that the tip of the siphon has a valve that regulates the outflow of water from the mantle cavity. The inner side of the mantle has cartilaginous ridges that keep the inhalent currents separate from the exhalent. There are two ctenidia (gills) oriented so that the inhalent streams pass over each, then converge and exit as a single exhalent stream.
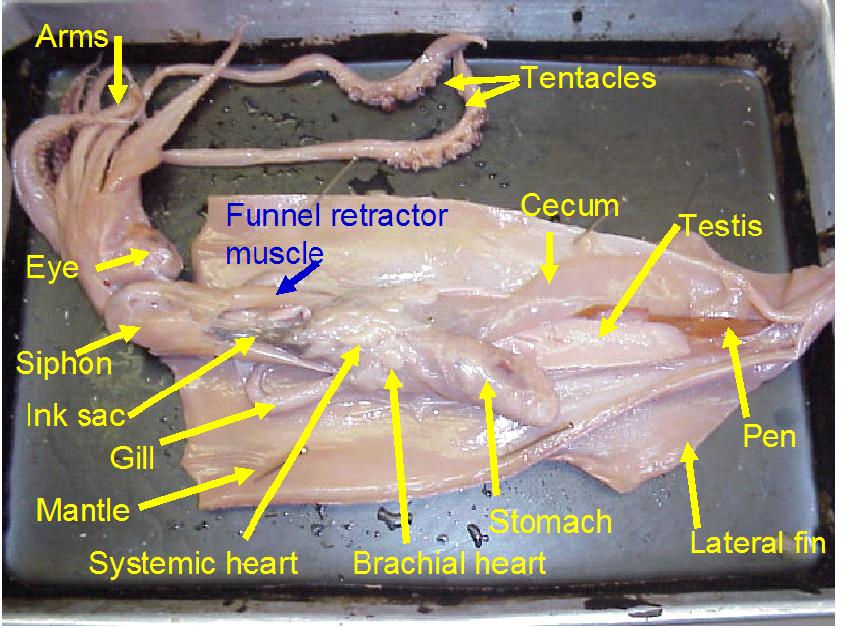
The paired excretory organs of the mollusks are closely associated with the heart. There are two kinds of molluscan excretory organs: brownish pericardial glands (difficult to see in most specimens) and renal nephridia or kidneys. In the kidney, nitrogenous wastes (urea, amines, and predominantly, ammonia) are extracted from the blood. Inorganic substances may be taken back into the blood, and the composition of the waste is determined by the extent of renal secretion and reabsorption. Excretory material is passed to the mantle cavity and voided in an excurrent blast through the siphon.
The cephalopod kidney is derived from two separate renal organs that have fused. The kidney is closely associated with the gill hearts, which are swellings of the blood vessels at the base of the gills. The contractions of the gill hearts forces fluid through the walls of the blood vessels into the kidney.
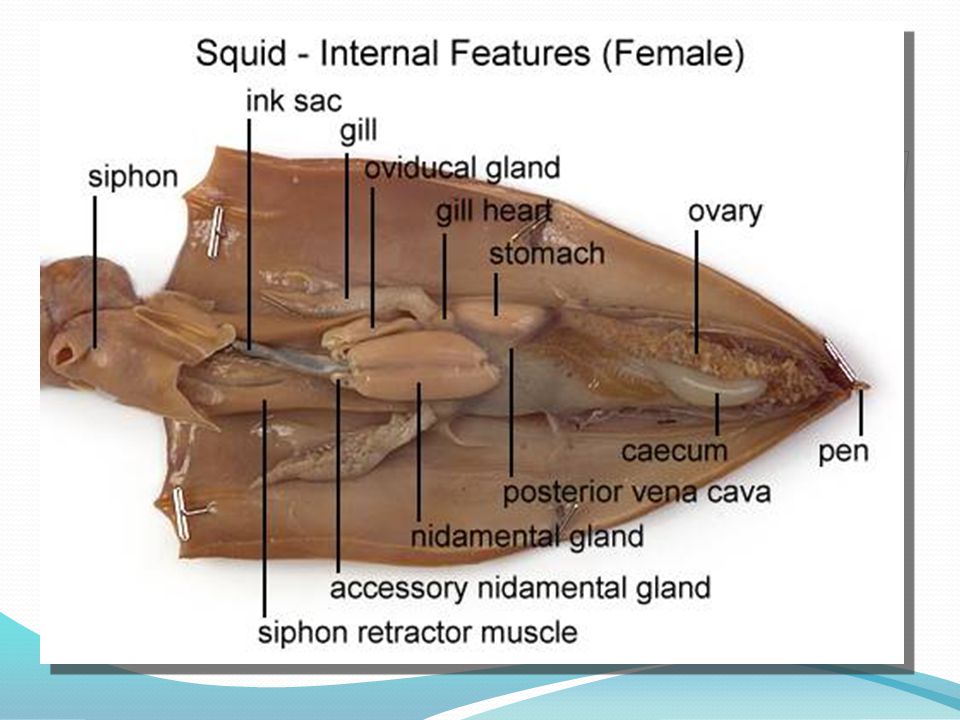
If necessary, in your opened Loligo, remove the thin semi transparent skin covering the organs of the visceral mass. Locate the rectum ending in the anus at the base of the siphon. If your specimen is a female, there will be two large white nidamental glands that covers most of the internal organs. These will need to be carefully removed in order to see the stomach and other organs. Notice the swollen branchial heart at the base of the left gill and attempt to look for it's close attachment to the kidney. This may be difficult to locate but give it a try.
Mollusks do not have a spacious body cavity (coelom), and in all except the cephalopods, the body space is composed of large sinuses, which act as pooling places for the blood. For this reason, the body space is more accurately described as a hemocoel; (a membranous sac enclosing the heart) and the cavity of the gonad. Blood that has collected in the sinuses from various parts of the body passes first through the kidney and respiratory organ and then via the branchial artery to the heart. The main propulsive force for distribution of blood in the connective tissue spaces gives the blood an additional function to that of distribution; namely, it acts as a component of the hydrostatic (fluid supported) skeleton. The foot of clams, for example, is enlarged and forced outward by the influx of blood and is withdrawn by contraction of longitudinal muscles.
The active swimming and carnivorous habits of the cephalopods require a more efficient circulatory system than in a clam - one that is closed and contains capillaries.
Mollusks demonstrate all possible feeding habits (herbivorous, carnivorous, and omnivorous), and the structure and function of the gut are related to the type of food eaten. Cephalopods are voracious carnivores and do not depend upon ciliary currents for the capture of food. They do, however, retain some of the structural components of the gut, such as the radula (a tooth-bearing, food-gathering structure unique to mollusks).
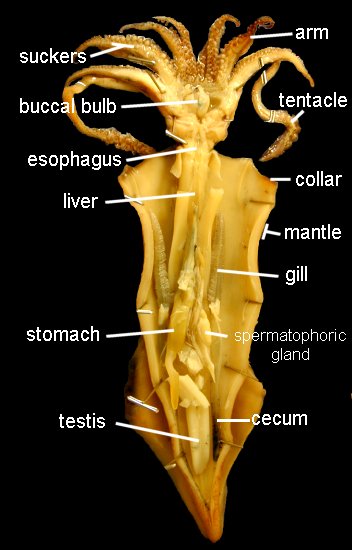
The mouth is surrounded by eight pointed arms and two longer tentacles. Observe the structure of these arms and tentacles and the arrangement of the suckers upon them. Study the organization of the suckers under the dissecting microscope. In life, the prey is grabbed by a rapid extension of the two tentacles and brought toward the mouth, where it is held firmly in place by the eight arms and killed by an injection of poison. The arms will then hold and manipulate the food while the squid devours its prey. Remove the siphon, and, by median incision, cut into the head, separating the eyes and exposing the buccal mass. This is a muscular organ that bears two horny beaks, which are used for ripping prey. Pry open the beak and observe the radula. Posterior to the buccal mass are a pair of salivary glands, which pour their poisonous secretions into the buccal cavity. Trace the thin-walled esophagus (surrounded by the liver) from the buccal mass to the thick-walled stomach. The stomach emerges from the liver tissue to form the caecum, which extends to the tip of the visceral mass. The liver consists of paired digestive glands fused in the midline; it is a triangular organ with the base located ventrally near the collar. A U-shaped pancreas lies anterior to the stomach; its duct unites with that of the liver before passing into the caecum. The intestine runs forward from the stomach, shows a diverticulum (the ink sac), and terminates in the rectum.
The structure of the nervous system in mollusks ranges from a simple "ladder" type to a system in which there is extreme fusion of ganglia, forming a true brain. The ultimate in invertebrate cephalization is seen in the cephalopods. Evolutionary fusion of ganglia followed by extensive differentiation makes it difficult to compare the ganglia with those of other mollusks. In your specimen, the easiest part of the nervous system to find are the large stellate ganglia, located on the inner dorsal surface of the mantle at the level of the tip of the ctenidia. The stellate ganglia are the motor centers of the mantle and give rise to its giant fiber system, which is favorite material for neurophysiologists.
Locomotion
From the complex structure of the cephalopod nervous system, one would expect these organisms to be capable of a variety of neuromuscular activities. They are particularly well demonstrated by the feeding and swimming movements of the squid. In Loligo, which is a rapid swimmer, the giant fiber system of the mantle is effective in producing quick-firing, rapid muscular contractions. In addition to moving quickly by use of the funnel system, when the squid feels threatened, it may release ink from its ink sac into the water as a way to confuse a larger predator. It will use this opportunity to then swiftly swim away to safety.
Vision
The eyes of cephalopods are the best developed among all mollusks, and perhaps among all invertebrates. Carefully remove an eye from your specimen by cutting the eye muscles and optic nerve. The outermost covering is the false cornea, which is underlain by the true cornea. Remove the corneas and identify the spherical lens, the colored iris and choroid coat, and the innermost lining, the retina. How does the organization of the eye of the squid compare with that of the human? In simple terms, the eyes of the squid are almost identical to the eyes of the human. Other than the false cornea, all of the other structures of the eye of the squid are the same as the structures in a human eye.
Reproduction
In cephalopods, the sexes are always separate; the gonad is at the apex of the body and its ducts open directly into the coelom. Fertilization is internal, and there may be complicated courtship behavior and parental care of the young. You should be sure to view the reproductive organs of both a male and a female squid.
If your specimen is a female, identify the single large ovary at the apex of the visceral mass. Identify the large, white nidamental glands, together with the orange-speckled accessory nidamental glands beneath their ventral ends. The oviduct is a transparent tube, possibly packed with eggs, leading by a small ciliated funnel from the vicinity of the ovary. The oviduct loops ventrally, dorsally, and ventrally again as the glandular, thicker walled oviducal gland. It terminates in a flared opening. The large, yolky eggs are shed from the ovary into the coelomic cavity and are picked up by the ciliated funnel of the oviduct. As the eggs pass along the oviduct, they receive an elastic membrane from the nidamental glands and a gelatinous coat from the oviducal glands.
If your specimen is a male, remove the left gill and branchial heart. The single large, white testis is located at the apex of the visceral mass. It opens directly to the coelom by a slit at its anterior end. Near the opening lies the ciliated opening to the vas deferens, an opaque, white, coiled tube leading ventrally between the spermatophoric sac on the right and the thick-walled spermatophoric organ on the left, and opening to the exterior by the penis to the left of the rectum. The penis is not a muscular intromittent organ and is no more than the end of the vas deferens.
Click HERE for the Squid Lab Companion (Identifies what material from the squid dissection will be on the lab exam.)